In Summer 2024, a team of palaeontologists and geologists from the University of Oxford, along with colleagues from Dartmouth College, the University of Washington, and Williams College in the USA, undertook an expedition to the Little Dal Group in the Mackenzie Mountains, Northwest Territories, Canada. Our purpose was to uncover some of the oldest fossil ecosystems that record complex life.
Photo: Robert GillPhoto: Robert Gill
Complex life comprises all organisms whose DNA is enclosed in a cell nucleus. This includes animals and plants but excludes bacteria. Today, this complex life accounts for most of the Earth’s biomass, documented biodiversity, and oxygen production. Understanding when and how it first evolved remains one of the central unanswered questions in evolutionary biology.
Photo: Robert GillPhoto: Robert Gill
As palaeontologists, we normally use fossils to reveal the history of life. Fossils tend to preserve larger animals with hard shells or skeletons—creatures such as trilobites, ammonites, dinosaurs, and mammoths. However, the first complex organisms were microscopic and lacked such hard parts. As a result, their soft and fragile cells rarely fossilised. Put simply, we have found it a major challenge to trace the origins of complex life with fossils.
Photo: Robert GillPhoto: Robert Gill
I have argued that finding rocks made up of antibacterial clay minerals holds the key. These minerals can slow the decay of organic cells long enough for them to survive as fossils. The Little Dal Group contains ~900-million-year-old rocks that are rich in just such clays, making it a prime target for new fossils that might help us unravel the origins of biological complexity.
Photo: Robert GillPhoto: Robert Gill
I was joined in Canada by my DPhil student, George Wedlake, from the Department of Earth Sciences. Together we spent two weeks collecting over 100 rock samples. The samples record an ancient tropical sea not unlike the Bahamas today, where early complex life likely flourished.
Back in Oxford, at the Museum of Natural History, George and I are now examining the samples; dissolving the rocks with hydrofluoric acid to extract and study the tiny fossils. We hope these new fossils will transform our understanding of how complex life first took hold on our planet.
Our fieldwork was funded by a Royal Society University Research Fellowship and by the Oxford NERC Environmental Science Doctoral Training Partnership. It was conducted under permit and with the support of the Sahtú Dene people.
Looking through the collections at OUMNH never gets boring, but sometimes a drawer will open up to reveal something even more eye-catching than the fossils usually found inside. Whilst working on the Museum’s Jurassic marine reptiles a few weeks ago, I came across something particularly surprising: a jewel-green box with a fantastic piece of art on the front. I was instantly intrigued and reminded of all the other times I had encountered a holder as fascinating as the specimen inside it.
Storage in museum collections is an ongoing pursuit of balance between ideal environmental conditions, specimen accessibility, and efficient use of space. This balance applies to all levels of storage: from building to room, cabinet to specimen tray. OUMNH’s Earth Collections are stored in conservation-grade, acid-free boxes or trays made of plastic or cardboard. These boxes are sometimes layered with low-density foam or ‘plastazote’ which can be carved to fit the specimen and keep it from being jostled or damaged. Holders with lids can also provide a micro-environment for specimens to help minimise their exposure to changes in humidity and temperature. The use of these standard materials not only helps protect specimens from degradation but can also deter pests from harbouring in collections spaces.
However, historical collections like those at OUMNH may retain holders that are not standard use. Sometimes, a clean and empty plastic Ferrero Rocher box is the perfect size for that small mammal skeleton that needs storing! Other times, an unusual holder might have been the only thing a field collector had on hand to transport a specimen to the Museum.
A harmonica box containing pliosaur teeth, a marine reptile that lived during the Jurassic (145.5 million – 201.6 million years ago).
One example of an unusual specimen holder is this ‘Echo Harp’ box by pre-eminent German harmonica manufacturer Hohner, likely from the 1960s. The box no longer holds a harmonica, but instead accompanies pieces of Jurassic pliosaur teeth from Weymouth, Dorset. Pliosaurs were a kind of carnivorous marine reptile related to plesiosaurs, with four flippers, and long tails and necks. If they hadn’t gone extinct in the Cretaceous-Paleogene extinction event 66 million years ago, perhaps they would have come to appreciate the harmonica and its artistic packaging!
Aside from their artistic value, museums may sometimes retain unusual holders because they contain primary source information on the specimen. One such example is a ‘Bryant and May’s Patent Safety Matches’ box in our Earth collections, bearing a packaging design from the early 1900s. The box actually houses a chicken tarsometatarsus bone excavated from “High St. New Schools” in Oxfordshire and is accompanied by a label which describes the particular layer of gravel the specimen was found in — important information for any archaeological or palaeontological find. Although the specimen is stored alongside Pleistocene fossils (10,000 – 2.6 million years ago), chickens did not originate in the UK, so the bone is likely from much more recent times. Someone still must have thought it was important enough to keep in its own special holder!
A Tate and Lyle sugar bag containing a Jurassic specimen, with handwriting on the outside describing the stratigraphy the fossil was found in.
Similarly, this ‘Tate and Lyle Granulated Sugar’ paper bag features a handwritten original notation in blue pen on the outside. The bag originally contained a specimen found in a collection of Jurassic gastropods and bivalves from Somerset, with the handwriting describing the fossil’s stratigraphic information. The bag also features a recipe for cinnamon apples on the reverse, which we have yet to try!
A wooden box and the Quarternary fossils (up to 2.6 million years ago) it originally housed. An accompanying letter describes the delivery of the fossils to William Buckland, Oxford University’s First Reader in Geology.
In addition to primary source information, original holders may also provide specimens with provenance. This ovular wooden box filled with organic stuffing material originally contained Quarternary fossil specimens found in Peak’s Hole, Derbyshire. The Museum archive also holds a handwritten letter describing the specimens inside the package and how they were found. The letter dates to 1841 and is addressed to Oxford University’s first Reader in Geology, William Buckland. The specimen holder forms part of a group of objects with such a strong interconnection, and such strong documentation, that retaining the box is a matter of course.
All in all, it’s great that we’ve come so far in the advancement of safe and stable housing for specimens. At the same time, it’s always fascinating to see what else has made its way into collections, just by nature of being able to hold things, either for a short time or a long one. Despite living in the Earth Collections – among fossils, rocks, and the geological past – these objects offer us a little bit of human history too.
UNEARTHING THE PECULIAR EATING HABITS OF A TRIASSIC MAYFLY SPECIES
During the summer months, the beaches of Mallorca offer an irresistible draw for tourists and palaeontologists alike. Visitors to the small Spanish island find themselves lured by its glittering seas, captivating coastline, and tasty white sands…
…well, tasty for some, at least!
Coastal cliffs near Estellencs (Mallorca, Spain). Palaeontologists working here discovered fossils of Triassic mayfly nymphs with unusual gut contents. (photo: Balearic Museum of Natural Sciences)
Following recent fossil excavations near the the coastal town of Estellencs in southwest Mallorca, palaeontologists have discovered evidence of a species of mayfly with a pretty peculiar diet. The mayflies in question lived 240 million years ago in bodies of water associated with ancient floodplains. Some of the juvenile mayflies (nymphs) were so well-fossilised that it has been possible to study the contents of their guts. A research team, led by Dr Enrique Peñalver, and featuring OUMNH’s own Dr Ricardo Pérez-de la Fuente, discovered that the mayflies’ digestive tracts contained a mixture of detritus (the decomposed remains of other organisms) and particles of a type of rock known as claystone. The most likely explanation for this strange food-pairing? It seems that the nymphs actually survived by eating muddy sediments that had settled to the bottom of the swampy-waters they lived in – yum!
If you’ve ever tried eating a sandwich on the beach, you’ll be familiar with the feeling of sand in your teeth. The sharp crunch of mineral sediment is worth the sacrifice for the delicious, digestible portion of your sandwich – the bread and fillings. Animal digestive systems are unable to extract energy from inorganic mineral matter, like sand. Instead, we rely on organic material for nutrition, i.e. matter derived from plants and other animals. It seems that the Triassic mayfly nymphs found in Mallorca would have munched through large quantities of sediment; digesting the organic detritus it contained, and excreting the inorganic remainder.
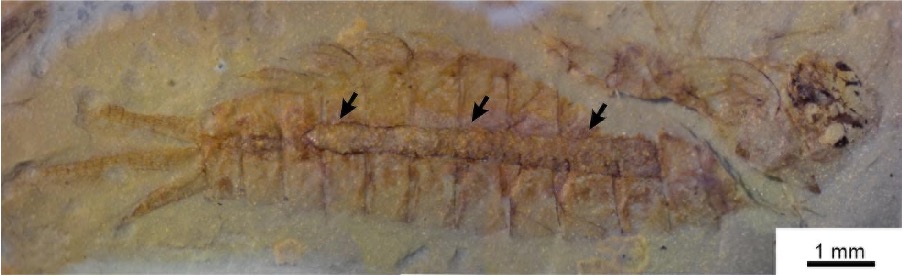
One of the numerous Early Triassic mayfly nymphs from Mallorca preserved with gut contents. These inclusions result from the original sediment the nymphs fed on (cololite, labelled here with arrows). Image adapted from Peñalver et al. (2023).
Sediment-based diets are extremely rare among living insect species. A handful of modern mayfly species have been observed to munch on the muddy sediment that surrounds the openings of their tunnels, but this is a very rare occurrence. Sediment is a pretty challenging food source, and it’s hard to say why insects may have relied more heavily on it in the ancient past. It is possible that the mayflies found in Mallorca adopted their diet as a result of the Permian mass extinction, which killed off more than 80% of all the species on Earth, ‘just’ five million years prior. With fewer choices of organic material available to eat, perhaps the mayflies were left without a better choice? Or maybe they were simply exploiting new environmental niches that opened up in the aftermath of this catastrophic event?
One of the reasons why it is so difficult to theorise about the evolution of species following the Permian mass extinction is the dearth of fossil evidence dating from the period. Luckily, the coastal cliffs of Mallorca can offer us a rare, exciting glimpse into some of the ecosystems that existed ~247 million years ago. The research team behind the Mallorcan mayfly discovery have also used fossils from the same site to describe the world’s oldest-known dipteran (a group of insects including flies, mosquitoes, gnats, and midges), naming the species Protoanisolarva juarezi. These flies would have lived on land, in back swamp areas, rather than in the water. However, much like the Triassic mayfly nymphs, they would have fed on detritus, and played a key role as recyclers of organic matter in these ancient ecosystems.
The larva of the oldest-known gnat, 247 million years old, was found near Estellencs in Mallorca. (Image: CN-IGME CSIC).
It is by paying attention to tiny insect fossils like these that we might hope to find answers to one of the biggest questions in palaeontology: how did life rebuild in the aftermath of our planet’s worst mass extinction? And what might this teach us about ecosystem responses to future mass extinction events?
By Ella McKelvey, Web Content and Communications Officer
If you’ve ever visited the Skeleton Parade in the Main Court of the Museum, you may have noticed that nestled between the Malayan tapir and the rhinoceros is the skeleton of a reindeer, or caribou if you are from North America.
Today, reindeer are found throughout the Arctic and Subarctic in places like Canada, Alaska, Russia, and Lapland (Norway, Sweden, and Finland). However, their range was not always so limited. During the Late Pleistocene – around 126,000 to 11,700 years ago – it would not have been unusual to see herds of reindeer roaming freely across most of Britain and western Europe. In fact, reindeer sub-fossils in the form of bones, teeth, and antlers have been found at a number of Oxfordshire sites including the excavations at Cassington and Sutton Courtenay, which are kept behind the scenes in the Museum’s extensive Paleontological Collections.
Studying these Ice Age reindeer can teach us as much about the future as they can about the past. Pleistocene reindeer were likely similar to their modern counterparts, which undertake large, bi-annual migrations between summer and winter grazing pastures. Looking at the movements of Ice Age populations of reindeer can therefore help us to understand how modern reindeer may respond to climactic and environmental changes in the future. This is possible because reindeer only come together in large herds at certain times of the year. During these seasonal aggregations, the herd is characterised by different combinations of ages and sexes. Therefore, by looking at the age and sex of the remains of reindeer present at a site, we can tell the time of year that they were left there — in particular, we can infer the sex of reindeer from their bones, their age from their teeth, and their age and sex from their antlers.
Modern reindeer are highly adapted to cold environments (-45 to +15°C) with two layers of fur (the tips of which turn white in the winter), short and furry ears and tails, and large feet to make walking on snow and digging for food much easier. Reindeer even make a clicking noise with their feet, produced by a tendon slipping over a bone, to help keep track of each other in blizzards or fog.
Unfortunately, it is extremely rare to find anything so complete as the reindeer in the skeleton parade. Instead, you are much more likely to find remains like the antler below, which was excavated from Sutton Courtenay. Despite being only a fragment, it is exactly this kind of sub-fossil that can help us to understand more about the movements of reindeer during the Late Pleistocene.
This left antler base and skull from a male reindeer found at Sutton Courtenay can be used to determine which season reindeer were present at the site.
Reindeer grow and shed a new pair of antlers every year, and this happens at different times of the year for males and females. If you can identify whether an antler is male or female, shed or unshed, you can also tell the season of death. The Sutton Courtenay antler featured above would have belonged to a male reindeer. At its base, we can see it is still clearly attached to some skull bone, and so is unshed. Because males only have their fully grown antlers between September and November, this particular reindeer must have been in the area around Sutton Courtenay during the autumn. It is by using similar deductions that we can also tell that Rudolph and his antlered friends would have actually all been females — by the 24th December, males have already shed their antlers, but females will keep them until the spring!
After studying thousands of these kinds of remains from all over Britain, we can start to build a picture of where reindeer were at different times of the year. It’s amazing to think that we can learn so much from simple skeletons. So, the next time you visit the skeleton parade, take a moment to think about the secrets they may be hiding.
By Chloe Williams, History Finalist at Oxford University and Museum Volunteer
Email: chloegrace1000@gmail.com
“The professor regrets to have to record the loss of the invaluable services of Miss Healey, who as a result of overwork has been recommended to rest for an indefinite period. This will prove a serious check to the rate of progress which has for some time been maintained in the work of rearrangement, and it is hoped that her retirement may be only temporary.” So ends the Oxford University Museum of Natural History’s 1906 Annual Report, marking the near-complete departure of Maud Healey from the archival record.
Despite how little of her history has been preserved, it is clear that Maud Healey made significant contributions to the field of geology. After studying Natural Sciences at Lady Margaret Hall in 1900, Healey worked at the Museum as an assistant to Professor William Sollas from 1902–1906. Here, she catalogued thousands of specimens and produced three publications. These publications were at the centre of debates about standardising the geological nomenclature, and turning geology into a practical academic discipline that could sustain links across continents. However, Healey was continually marginalized on the basis of her gender. Closing the Geological Society of London’s discussion of one of her papers, “Prof. Sollas remarked that he had listened with great pleasure to the complimentary remarks on the work of the Authoress, and regretted that she was not present to defend before the Society her own position in the disputed matter of nomenclature.”[1] Predating the Society’s 1904 decision to admit women to meetings if introduced by fellows, Healey had been unable to attend the reading of her own paper.
Photo of the Geological Society of London centenary dinner in 1907, at which Maud Healey was present. Healey can be seen seated in the fourth row from the front, three chairs to the left. Of the 263 guests, 34 were women, 20 of whom were the wives or daughters of academics, and only 9, including Healey, were present ‘in their own right’. [2] Source: Burek, Cynthia V. “The first female Fellows and the status of women in the Geological Society of London.” Geological Society, London, Special Publications 317, no. 1 (2009): 373-407.
Healey later worked with specimens collected by Henry Digges La Touche in colonial Burma (now Myanmar). While Healey worked with the identification of species, acknowledged by La Touche himself as ‘a more difficult lot to work at’ than similar specimens assigned to her male contemporaries, the physical collection and therefore its name and record is attributed to a male geologist. [3] She continued her work identifying La Touche’s collection of Burmese fossils after retiring from the Museum in 1906 and published a report about them in 1908. What happened to her afterwards is unclear. Tantalizing snippets like a 1910 marriage record might suggest that she turned to a life of domesticity, but whether Healey continued to engage with geology as a hobby remains uncertain.
Left: Fossil bivalves from Burma (Myanmar), part of the La Touche collection sorted and identified by Maud Healey. Right: Illustration from Fauna of the Napeng Beds (1908), Maud Healey’s monograph describing the La Touche collection. From the library at Oxford University Museum of Natural History.
It is almost unbelievable that a professional of Healey’s calibre could abandon the work in which she excelled. However, Healey lacked any familial connections to geology, and apparently did not marry into money, which would have made it difficult for her to retain access to organizations like the Geological Society of London. The diagnosis of ‘overwork’ mentioned in the Annual Report makes it possible that a medical professional could have discouraged her from engaging further in academia. Unfortunately, any diaries or letters which might have provided us with further clues were not deemed worthy of preservation.
Maud Healey on a dig site (location unknown). Image from the Archives at Oxford University Museum of Natural History.
Tracing Maud Healey’s history to 1910, it might seem as though we hit a depressing dead end. Healey is one of many nineteenth-century female geologists who participated in an international community in a range of roles including collecting, preserving samples, and actively producing knowledge. However, like many of her colleagues, her contributions are largely absent from the historical record. My research doesn’t aim to simply ‘rediscover’ these exemplary women after previously being ‘hidden’ from history, but instead considers how history itself is constructed from a material archive created along lines of gender and class. A subjectivity which surfaces only rarely in appended discussions to academic papers, and in spidery cursive on ancient fossils, Maud Healey ultimately suggests the need for women’s history to read archival silences as their own stories.
Works cited
[1] Healey, M. ‘Notes on Upper Jurassic Ammonites, with Special Reference to Specimens in the University Museum, Oxford: No. I’, Quarterly Journal of the Geological Society of London 60, (1904), p.1-4.
[2] Burek, Cynthia V. “The first female Fellows and the status of women in the Geological Society of London.” Geological Society, London, Special Publications 317, no. 1 (2009): 373-407.
[3] La Touche, H.D. Letter to Anna La Touche, 1 August 1907. La Touche Collection. MSS.Eur.C.258/77. Asian and African Studies Archive, The British Library, London, UK.
Susan Newell is a doctoral student researching the teaching collections of William Buckland, the first Professor of Geology at Oxford who taught from 1813 to 1849. She reminds us here about Buckland’s role 200 years ago in interpreting the important Pleistocene discoveries being celebrated this year, and the way that Mary Morland, a talented local naturalist, and many others, contributed to making this new knowledge.
This year marks the 200th anniversary of a great advance in our understanding of the geological past… a story which begins in the nineteenth century, with the discovery of a bone-filled cave in Kirkdale, Yorkshire.
Uncovered by local quarrymen in 1821, the discovery of the Kirkdale cave and its contents of mysterious bone was the source of much intrigue. When news of the discovery reached William Buckland, Professor of Geology at Oxford University, he decided to travel up North to visit the site. However, by the time Buckland arrived at the cave, local collectors had scooped up most of its contents. Nonetheless, he was able to retrieve and examine some of the cave’s remaining material, which led him to an astonishing conclusion — Yorkshire must once have been home to hyaenas, elephants, hippopotamus and rhinoceros, and what was now known as the Kirkdale cave was once a hyaenas’ den.
W. B. Conybeare, lithograph, ‘The Hyaena’s Den at Kirkdale near Kirby Moorside in Yorkshire, discovered A.D. 1821’. Reproduced by kind permission of Christ Church, Oxford. This light-hearted reconstruction of the hyaenas’ den shows Buckland illuminating the scene, in every sense. It is thought to be the first visual reconstruction of the pre-human past.
Central to Buckland’s theories were some small white balls that he had found amongst the debris in the cave. Buckland sent these balls to William Wollaston, a celebrated chemist based in London, for analysis. He also asked Wollaston to visit the zoo at Exeter Exchange in London and show the balls to the hyaena’s keeper there. Together with the results from Wollaston’s chemical analyses, the keeper confirmed Buckland’s hypothesis — the balls were droppings from animals very similar to modern hyaenas. Meanwhile, the anatomist William Clift was able to identify the bones from the Kirkdale cave as belonging to other extinct species related to those found living in tropical countries today. Buckland concluded that the cave must have been a den for ancient hyaenas, who would drag parts of the dead animals they had found (or killed) inside and, after feeding on them, leave piles of bones and droppings behind.
In order to strengthen his theory, Buckland discussed the behaviour of hyaenas in the wild with army officers connected to Britain’s colonial expansion in India. These officers also sent Buckland fresh specimens captured by local people. When a travelling menagerie visited Oxford in 1822, Buckland took the opportunity to experiment; feeding bones to a hyaena and noting that the teeth marks matched those on the fossilised bones from the cave.
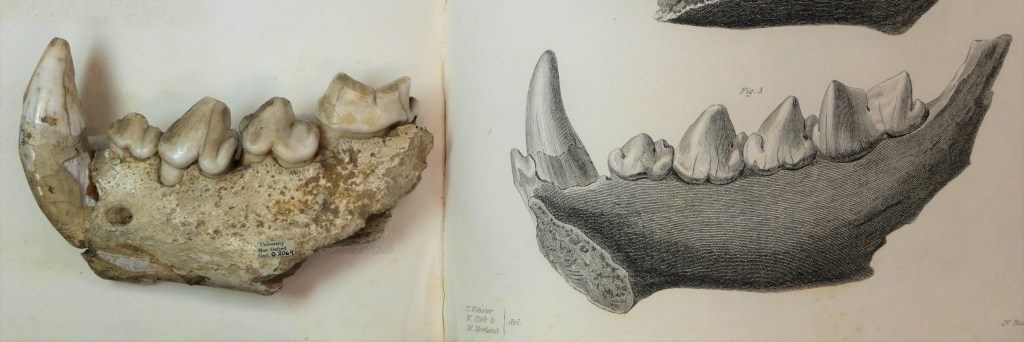
Left: “Small white balls” of hyaena excrement. The left box probably contains samples collected from the hyaena in the travelling menagerie which visited Oxford, and the right contains fossilized samples collected by Buckland from Kirkdale Cave, now part of the Museum’s Collections. Right: Comparison of two shinbones – one fed to a hyaena by Buckland, and the other collected from Kirkdale cave. The similarity of the gnaw-marks suggest that hyaenas were once present in Kirkdale. On display in the Museum.
Buckland’s findings were something of a shock to his contemporaries. When lecturing, he employed several different methods to try and convince his audiences that his theories were true. This included presenting fossil specimens and bones from living species for comparison, and showing maps, diagrams and drawings. Mary Morland contributed some of these illustrations, including large drawings of living animals, and technical drawings of bones that were later engraved for use in Buckland’s publications. Mary’s Kirkdale drawings seem to have been the first that she produced for William before the couple married in 1825.
Fossil hyaena jaw in the Museum’s collection, possibly the one featured in the engraving alongside it. Engraving is by James Basire after a drawing by Mary Morland. Published in William Buckland’s article in the Royal Society’s journal (1822) on the Kirkdale cave discoveries. [1]
Buckland’s work on the Kirkdale cave was revolutionary, not least because he was the first to make a scientific study of a cache of bones of this type. Although similar bones from ‘tropical’ species had previously been found in Northern Europe, people thought that they had been washed up by a catastrophic flood, believed by many to be the biblical Noah’s Flood. Modern analysis has now allowed us to deduce that the bones date to an Interglacial period when Britain was joined to Europe and had a hot climate, about 120,000 years ago.
Here at the Museum, Buckland’s collections and archives are as much of a treasure trove as the Kirkdale cave. It is through accessing these archives that we can learn about the surprising range of people who contributed to the emergence of new scientific knowledge from the Kirkland cave — quarrymen, collectors, zookeepers, chemists, anatomists, colonial officers in India, workers in India, and artists like Mary Morland. To find out more about the incredible legacy of the Kirkdale Cave, look out for ‘Kirkdale200 – Lost Beasts of the North’, a symposium organised by the Yorkshire Fossil Festival, 12th March 2022.
Mary Morland, watercolour and gouache, lecture illustration of a hippopotamus, signed ‘MM’. Hippopotamus bones were found at Kirkdale cave in Yorkshire, but as there were no living hippos to be seen in Britain at the time, this drawing would have been a valuable teaching aid.
[1] William Buckland, ‘Account of an Assemblage of Fossil Teeth and Bones of Elephant, Rhinoceros, Hippopotamus, Bear, Tiger, and Hyaena, and Sixteen Other Animals; Discovered in a Cave at Kirkdale, Yorkshire, in the Year 1821: With a Comparative View of Five Similar Caverns in Various Parts of England, and others on the Continent’, Phil. Trans., 2 (1815-30), 165-167.